Sponges have several cell types:
* Choanocytes (also known as "collar cells"), flagellated cells which function as the sponge's digestive system, are remarkably similar to the protistan choanoflagellates. The collars are composed of microvilli and are used to filter particles out of the water. The beating of the choanocytes’ flagella creates the sponge’s water current.
 * Porocytes are tubular cells that make up the pores into the sponge body through the mesohyl. * Porocytes are tubular cells that make up the pores into the sponge body through the mesohyl.
* Pinacocytes which form the pinacoderm, the outer epidermal layer of cells. This is the closest approach to true tissue in sponges.
* Myocytes are modified pinacocytes which control the size of the osculum and pore openings and thus the water flow.
* Archaeocytes (or amoebocytes) have many functions; they are totipotent cells which can transform into sclerocytes, spongocytes, or collencytes. They also have a role in nutrient transport and sexual reproduction.
* Sclerocytes secrete calcareous or siliceous spicules which reside in the mesohyl.
* Spongocytes secrete spongin, collagen-like fibers which make up the mesohyl.
* Collencytes secrete collagen.
* Spicules are stiffened rods or spikes made of calcium carbonate or silica which are used for structure and defense.
* Cells are arranged in a gelatinous non-cellular matrix called mesohyl.
Body types: Sponges have three body types: asconoid, syconoid, and leuconoid.
Asconoid sponges are tubular with a central shaft called the spongocoel. The beating of choanocyte flagella force water into the spongocoel through pores in the body wall. Choanocytes line the spongocoel and filter nutrients out of the water.
Syconoid sponges are similar to asconoids. They have a tubular body with a single osculum, but the body wall is thicker and more complex than that of asconoids and contains choanocyte-lined radial canals that empty into the spongocoel. Water enters through a large number of dermal ostia into incurrent canals and then filters through tiny openings called prosopyles into the radial canals. There food is ingested by the choanocytes. Syconoids do not usually form highly branched colonies as asconoids do. During their development, syconoid sponges pass through an asconoid stage.
Leuconoid sponges lack a spongocoel and instead have flagellated chambers, containing choanocytes, which are led to and out of via canals.
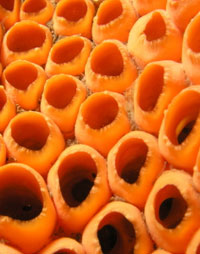 Physiology: Sponges have no real circulatory system, however the water current is used for circulation. Dissolved gases are brought to cells and enter the cells via simple diffusion. Metabolic wastes are also transferred to the water through diffusion. Physiology: Sponges have no real circulatory system, however the water current is used for circulation. Dissolved gases are brought to cells and enter the cells via simple diffusion. Metabolic wastes are also transferred to the water through diffusion.
Pumping: Sponges pump remarkable amounts of water. Leuconia, for example, is a small leuconoid sponge about 10 cm tall and 1 cm in diameter. It is estimated that water enters through more than 80,000 incurrent canals at a speed of 6cm per minute. However, because Leuconia has more than 2 million flagellated chambers whose combined diameter is much greater than that of the canals, water flow through chambers slows to 3.6cm per hour. Such a flow rate allows easy food capture by the collar cells. All water is expelled through a single osculum at a velocity of about 8.5 cm/second: a jet force capable of carrying waste products some distance away from the sponge.
Excretion and respiration: Sponges have no respiratory or excretory organs; both functions occur by diffusion in individual cells. Contractile vacuoles are found in archaeocytes and choanocytes of freshwater sponges. The only visible activities and responses in sponges, other than propulsion of water, are slight alterations in shape and closing and opening of incurrent and excurrent pores, and these movements are very slow.

 Both ways: Sponges are able to reproduce sexually or asexually. Most sponges are monoecious. Sperm are formed from choanocytes; and oocytes derive from choanocytes in some sponges, and archaeocytes in others. Sperm are released into open water. Some sponges have oviparous fertilization where sperm and eggs meet in open water. In other species sperm are taken into the bodies of other sponges and fertilized inside; this is known as viviparous fertilization. The free-swimming larvae, known as parenchymula, serve as one of the main forms of dispersal for this largely sessile phylum. Both ways: Sponges are able to reproduce sexually or asexually. Most sponges are monoecious. Sperm are formed from choanocytes; and oocytes derive from choanocytes in some sponges, and archaeocytes in others. Sperm are released into open water. Some sponges have oviparous fertilization where sperm and eggs meet in open water. In other species sperm are taken into the bodies of other sponges and fertilized inside; this is known as viviparous fertilization. The free-swimming larvae, known as parenchymula, serve as one of the main forms of dispersal for this largely sessile phylum.
Asexual reproduction: Asexual reproduction of sponges is through budding, where a small piece of sponge falls off of the main sponge and grows into a new one. Many freshwater sponges form small structures known as gemmules, which function as overwintering devices. These are similar to a bacterium's endospore. Gemmules are made up of amoebocytes surrounded by a layer of spicules and can survive conditions that would kill adult sponges. When the environment becomes less hostile, the gemmule resumes growing.

There are over 5,000 modern species of sponges known, and they can be found attached to surfaces anywhere from the intertidal zone to as deep as 8,500 m (29,000 feet) or further. Though the fossil record of sponges dates back to the Precambrian era, new species are still commonly discovered.
 Primitive: Sponges represent the most primitive animals, and as such, lack many of the typical features of animals including nerves and locomotion. Because of this they are placed in the group Parazoa, which is a sister taxon to all higher animals (metazoa). Sponges share many characteristics with colonial protists, such as Volvox, though they have a higher degree of cell specialization and interdependence. However, if a sponge is placed in a blender, any surviving cells can reform a complete organism. If multiple sponges are blended together, each species will recombine independently (contrast animal chimera such as the geep). Primitive: Sponges represent the most primitive animals, and as such, lack many of the typical features of animals including nerves and locomotion. Because of this they are placed in the group Parazoa, which is a sister taxon to all higher animals (metazoa). Sponges share many characteristics with colonial protists, such as Volvox, though they have a higher degree of cell specialization and interdependence. However, if a sponge is placed in a blender, any surviving cells can reform a complete organism. If multiple sponges are blended together, each species will recombine independently (contrast animal chimera such as the geep).
Sponges are divided into classes based on the type of spicules in their skeleton. The three classes of sponges are bony (Calcarea), glass (Hexactenellida), and spongin (Demospongiae). Some taxonomists have suggested a fourth class, Sclerospongiae, of coralline sponges, but the modern consensus is that coralline sponges have arisen several times and are not closely related.[1] In addition to these three, a fourth class has been proposed: Archaeocyatha. While these ancient animals have been phylogenetically vague for years, the current general consensus is that they were a type of sponge.
Although 90% of modern sponges are demosponges, fossilized remains of this type are less common than those of other types because their skeletons are composed of relatively soft spongin that does not fossilize well. The fossil Archaeocyantha may also belong here, though their skeletons are solid rather than separated into spicules. It has been suggested that the sponges are paraphyletic to the other animals. Otherwise they are sometimes treated as their own subkingdom, the Parazoa. Similar fossil animals known as Chancelloria are no longer regarded as sponges. Sponges have holes throughout their bodies in addition.
One phylogenetic hypothesis based on molecular analysis proposes that the phylum Porifera is in fact paraphyletic, and that members of Porifera should be split into two new phyla, the Calcarea and the Silicarea.
All text is available under the terms
of the GNU Free Documentation License
|